1 、简介
呼吸细胞中普遍存在一种奇特的酶活性,底物是一种能在任何瞬间微量存在的不稳定自由基,即使在此酶缺失的情况下,这种催化反应也能快速进行。然而这种酶对于需氧细胞的存活是至关重要的,它可以催化清除超氧自由基,因而可以抵御氧中毒,因为超氧自由基是产生氧中毒的重要物质。超氧自由基清除反应是歧化反应,这种酶可提高反应速率,反应式如下。

2 、超氧自由基和歧化反应
O2- 是氧还原的一种常规媒介,这是氧分子在基态时更倾向于单价还原途径的结果。产生O2-的反应很多,如对苯二酚、无色核黄素、儿茶酚胺、硫醇、还原染料、四氢蝶啶、铁氧化还原蛋白、红氧化还原蛋白和血红素蛋白等的自氧化反应。此外,很多酶促反应也可产生O2-,包括黄嘌呤氧化酶、醛氧化酶、二氢乳清酸脱氢酶及一系列黄素蛋白脱氢酶。很多氧化酶和羟化酶可被超氧化物歧化酶(SOD)抑制,这显示O2- 是这些酶催化循环的媒介,如色氨酸二加氧酶、重建肝微粒体羟化酶、黑曲霉源可溶性羟化酶和半乳糖氧化酶。
最终,O2-可由完整粒细胞吞噬时产生或由叶绿体接受光照时产生,也可由冻干的大鼠肝微粒体产生。可以肯定的是呼吸细胞能够产生大量O2-。超氧自由基不能在水介质中积聚,因为它极易发生歧化反应。O2-是过氧羟自由基弱酸的共轭碱,该弱酸的pKa值是4.8。自发歧化反应对pH的依赖性可以通过pKa和下列反应得到解释。

pH 7.4时,自发歧化反应的速率常数约为2×10(5次方) (mol/L)-1·s-1。在此pH下,若要通过SOD的作用加快O2- 的清除,取决于两个因素。第一,O2- 和SOD的反应速率常数应该接近2×10(9次方) (mol/L)- 1·s-1,使该催化反应比pH 7.4时的自发歧化反应快104倍。第二,细胞内酶的浓度要大大超过O2- 的稳态浓度,才能使得超氧自由基与酶的碰撞速率远远大于与另一O2- 的碰撞速率。据估计人类肝脏内SOD的浓度至少为3×10(负5次方) mol/L,而O2-的稳态浓度必须比SOD的浓度至少低5个数量级,使得O2- 的酶催化清除反应至少加快105倍。这两个因素的结合,将使肝脏内O2- 在pH 7.4时的酶催化清除反应至少比自发清除反应快10(9次方)倍。
3 、SOD的活性分析
对于酶的生产研究,其首要要求是快速准确的检测方法。对SOD而言,由于其底物不稳定,要达到这一目标需要费些周折。将产生O2-的反应和O2-指示清除剂联合在一起是一种成功的策略。在这种情况下,SOD与指示清除剂竞争O2-,从而抑制了清除剂与O2-的反应。有氧条件下,黄嘌呤氧化酶作用于黄嘌呤会产生O2-,该反应可以通过O2-对细胞色素c的还原能力检测到。还原型细胞色素c在550 nm有最大吸收,而SOD存在时会抑制细胞色素c的还原反应。SOD的一个活性单位可定义为在特定条件下抑制细胞色素c 还原率达50 %的酶量。
在SOD的首次分离中,该分析方法的灵敏度为一个活性单位相当于0.1 μg/mL的酶。此灵敏度还可通过提高pH和降低细胞色素c的浓度而被大幅提高。O2- 可通过不同于黄嘌呤氧化酶反应的其他反应产生,O2-指示清除剂也可以是不同于细胞色素c的其他物质。SOD的简单测定可以根据其抑制自由基链式氧化反应的能力实现,其中O2- 是引发剂或链增长自由基。多种反应已用于这种测定方法,包括亚硫酸盐、肾上腺素和连苯三酚的自氧化反应。根据抑制直观过程的能力检测SOD的直接法是不常用的。该法中,SOD可加快底物消耗或产物生成,理论上比上述间接方法更好。
通过电子顺磁共振(EPR)波谱法或直接紫外分光光度法来检测O2- 的消耗量,以此直接测定SOD活性。紫外分光光度法中,利用脉冲辐射分解法可使含氧水溶液中快速产生高浓度O2-,然后直接测定其紫外吸收。以上研究或类似的研究对于探索SOD的作用机制是非常重要的。
4 、SOD的生物学重要性
SOD对呼吸细胞的存活是至关重要的。
(1)可能只有呼吸细胞产生O2-,因此只有这种细胞需要防御自由基。对大量微生物的调查表明,很多需氧和耐氧生物均含有SOD,而专性厌氧微生物则不含SOD。不含SOD的耐氧生物是不能利用氧气的。
(2)SOD在粪链球菌、大肠杆菌B和酿酒酵母中可被氧诱导,研究证实有高含量、氧诱导SOD的细胞可以抵抗高压氧。而对照实验枯草芽孢杆菌和大肠杆菌K12中,氧诱导的是过氧化氢酶,因此细胞在有氧条件下均不能抵御高压氧。
(3)大肠杆菌突变株K12C-600对SOD有温度敏感性缺陷,同时在耐氧性上也存在相同缺陷。但发现,其回复子能使限制性温度下的耐氧性复得,也能在此温度下维持正常细胞水平的SOD。
(4)链黑菌素可以作为O2- 的细胞内供应源,氧可提高链黑菌素对大肠杆菌的致死率。检测证明,氧诱导SOD可对该抗生素提供耐受性。微生物研究结果表明,SOD对氧和过氧化物的毒性防御至关重要。数据表明,对于高等生物也是这样。
(5)在黄素单核苷酸(FMN)和EDTA存在下培养胎牛成肌细胞,若将其暴露于光线下细胞会受损伤,培养基中添加SOD时可保护细胞,降低O2- 光化学通量对细胞的致死率。SOD可抵御蒽、甲基胆蒽或苯并芘由于增强自由基光化学通量而导致的细胞毒作用。还有报告指出SOD可防御γ射线的有害辐射。
(6)大鼠暴露于85 %氧的环境中可以慢慢适应,继而可耐受100 %的氧,这表明缺乏适应性时的致命性。肺SOD的适度重复性增加,也显示出同样的适应性。
(7)SOD可预防谷胱甘肽诱导的大鼠肝线粒体肿胀。
(8)不论何种给药途径,服用百草枯都会造成大鼠的肺部损伤,频繁间歇性注射SOD可降低百草枯的毒性。
5、 SOD的多样性
应激反应的发生,很可能迫使存活物种间做出相似而独立的适应性调整。蓝藻类的光合活性产生了地球大气层的氧合作用,造成了这种氧化应激,使得不同种类的SOD成为一种恰当的适应性调节剂,存在于物种中。
5.1 Cu, Zn-SOD
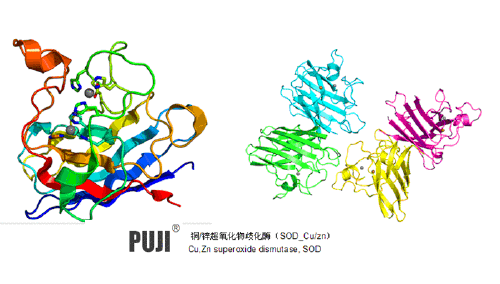
真核细胞的细胞质含有一种SOD,分子量为32 000,由2个亚基组成,每个亚基含有1个Cu2+和1个Zn2+。这种SOD具有明显的进化上的保守性,从真菌、植物、鸟类和哺乳动物中获得的酶非常难区分,只是在氨基酸组成和EPR波谱的超精细细节上存在相对较小的差异。这个酶曾可逆地分解。单独Cu2+复位可使活性部分恢复,两种金属离子都复位则可使活性全部恢复。试图使用其他金属取代Cu2+的实验无一成功,但Zn2+可以用Co2+、Hg2+或Cd2+替代,且活性不降低。牛红细胞SOD的结构分析正处于很理想的状态,得到了氨基酸的完整序列,且X射线衍射分析的分辨率已达到0.30 nm。
因此,建立了SOD的结构模型,并标明了每个氨基酸残基及Cu2+和Zn2+的位置。SOD最突出的结构特征是圆桶状结构,其外壁由八股序列以反平行β折叠的形式组成。β圆桶的序列片段为残基2-11,13-23,26-35,38-47,80-88,91-100,112-118和142-149。Cu2+ 和Zn2+ 通过His 61咪唑环配体加入进来,Cu2+ 的位置相对暴露于溶剂,而Zn2+埋藏于分子内部。除了共享的His 61,配位到Cu2+上的还有His 44,46和118,配位到Zn2+上的还有His 78和69及Asp81。
SOD的详细结构将促进对其机制的进一步研究。Cu2+和Zn2+ 的邻近性据推测是一种由组氨酸组成的Cu2+配位场。最初用脉冲辐射分解法探讨了SOD的这一机制,而且普遍认为,Cu2+ 在与O2-连续碰撞过程中被交替性地还原和再氧化。如果直接电子转移发生在O2- 和Cu2+ 之间,则Cu2+必须很容易进入水中。从序列分析和X射线衍射分析推论出的结构也证明了这种情况。NMR结果也表明Cu2+ 是暴露于溶剂的。氰化物与Cu2+ 结合时,其碳原子立即与Cu2+配位结合,因此氰化物是Cu, Zn-SOD的可逆抑制剂。另外,探讨了SOD分子中Cu2+ 的氧化还原性质。H2O2可以还原Cu2+,但浓度超过10 μmol/L时,也可以使酶不可逆失活。H2O2和Cu2+ 之间的氧化还原反应一定是单价的,因为SOD反应的可逆性已经被证实。
5.2 Mn-SOD

大肠杆菌B和变形链球菌含有的SOD,除了催化活性,与真核细胞质中的Cu, Zn-SOD显示毫无关联。这种SOD的每个亚基含有一个锰原子,分子量是40 000,是由大小相等的2个亚基组成的二聚体。一种提取自发光蘑菇Pleurotus olearius的Mn-SOD,也由大小相等的亚基组成,可在0.7 mol/L尿素存在时用离子交换色谱分离。
上述大肠杆菌SOD的颜色和NMR性质表明,它含有的锰是三价的。这种细菌性SOD的作用机制似乎和Cu, Zn-SOD相似,很可能取决于活性金属离子交替性地还原和再氧化。作为pH的函数,Mn-SOD的催化活性可用动力学竞争法进行研究。pH 7.0时,Mn-SOD的活性与Cu, Zn-SOD相似,pH升高时活性越来越低,而Cu, Zn-SOD的活性在pH 5.5~10.0范围内不受影响。线粒体中的一种SOD与原核生物中的Mn-SOD非常相似,不同的是线粒体SOD含有4个亚基而不是2个,且分子量接近80 000。
大肠杆菌和鸡肝线粒体中的Mn-SOD基本性质的相似性表明,它们有很近的亲缘关系。已经获得的部分氨基酸序列,也证明了线粒体SOD和细菌SOD间的紧密关系,但和细胞质SOD没有关联。这一结果为线粒体是起源于内共生于真核生物细胞中的原核生物的理论提供了强有力支持。最近,Mn-SOD已经从人肝线粒体中分离出来,与鸡肝线粒体SOD非常相似。牛肝中的高分子量SOD和人类多种器官中的同工酶B,都是上面提到的线粒体Mn-SOD。从Pleurotus olearius中提取的Mn-SOD很可能也存在于真核生物线粒体中,因为它与其他迄今为止研究过的线粒体Mn-SOD有很多相同的特性。
5.3 Fe-SOD

大肠杆菌B含有两种SOD,其中一个是Mn-SOD,另一个是位于细胞周质间隙的Fe-SOD,后者会因渗透压休克而被选择性地移出细胞。Fe-SOD的基本特性和氨基酸序列表明,它与来自大肠杆菌基质和鸡肝线粒体的Mn-SOD有较近的亲缘关系。因为氧合作用大肠杆菌中SOD含量的增加,其实是源于Mn-SOD的增加。由于这与高压氧耐受性的增加有关,由此可得出基质酶能清除内源性O2-的结论。细胞周质Fe-SOD的含量可以通过改变营养而改变,表明它可以防御外源性O2-。
近来认为,O2-对多形核白细胞有杀伤作用。与该观点一致的是,大肠杆菌B生长在富铁介质中时,其周质SOD含量会增加,而生长在缺铁介质中时,其Fe-SOD的含量会降低,因此富铁大肠杆菌与缺铁大肠杆菌相比能够更好的抵御吞噬细胞死亡。通过EPR和NMR分析,Fe-SOD中的铁离子是三价。在两类蓝藻[织线藻(Plectonema boryanum)和钝顶螺旋藻(Spirulina platensis)]和两类海洋细菌[乌贼发光杆菌(Photobacterium sepia)和鳆发光杆菌(Photobacteriumleiognathi)]中也发现了类似Fe-SOD。
6 、争议
有报道称牛红细胞SOD的亚基是共价键连接,这一结论是基于该亚基在没有巯基乙醇的情况下不能在十二烷基硫酸钠(SDS)中解离得出的。后来发现,此酶不能在SDS中解离不是因为其亚基是二硫键连接,而是因为连接键太稳定以致不能被SDS断开。事实上,它在1 %SDS中十分活跃,可以通过在SDS中加热或在SDS中加入尿素而被解离。在巯基乙醇中加入SDS才能解离,是因为链内的二硫键对全酶结构有稳定作用。据报道,SOD真正的生物功能是清除单态氧。
由于多个实例证明SOD能抑制大概是源于单态氧的化学发光,所以提议用缩写SOD(译者注:译文中此前及此后的SOD,在原著中作者均用superoxide dismutase或enzyme表示),即超氧化物歧化酶来代表单态氧清除酶(singlet oxygendecontaminase)。此外,单态氧可快速被羟基溶剂淬灭,在水中的寿命只有1 × 10-6 s。鉴于单态氧的寿命之短和能够有效猝灭化学发光反应的SOD的浓度之低,可以计算出SOD淬灭单态氧的速度应该至少高于其扩散速度5个数量级。

因此,SOD淬灭单态氧的报道在理论上就存在质疑,这也是SOD不能淬灭单态氧的直接证据。报道中也明确指出,SOD对单态氧的淬灭在很多方面存在缺陷。这种淬灭取决于单态氧只能是由过氧化铬(Ⅴ)酸钾在水中分解而来,但这种分解同时也释放O2-。此外,检测的是鲁米诺的化学发光反应,该反应在缓冲溶液中无论何种氧化剂总是依赖O2-,因此总是被SOD抑制。由此得出结论,SOD不能催化清除单态氧。真核细胞(如粗糙链孢菌和酿酒酵母)胞质、菠菜、牛和人类中的Cu, Zn-SOD都非常相似,但与原核生物或线粒体中的Fe-或Mn-SOD完全不同。氨基酸序列表明,细菌SOD和线粒体SOD的亲缘关系很近,但与真核细胞胞质SOD没有关联。这些结果具有前面讨论过的进化上的意义。
7 总结和展望
O2- 可作为氧化剂氧化肾上腺素、邻苯二酚和脱氢酶-NADH,且在肾上腺素和连苯三酚的自氧化反应中可作为链传递物质。O2- 也可作为还原剂,细胞色素c的还原是SOD活性测定的基础。O2-还可迅速还原铁蛋白中的Fe (Ⅲ)。在其他更复杂的系统中,O2-也表现有活性。SOD可防止谷胱甘肽引起线粒体肿胀,可保护细胞膜被X射线照射时的脂质过氧化,还可防止线粒体内膜在谷胱甘肽有氧溶液中培养时被过氧化。
另有报道也指出SOD可阻止脂质过氧化。5-羟巴比妥酸,自氧化时产生O2-,SOD可防止5-羟巴比妥酸造成的维生素E缺乏性大鼠红细胞的溶血。SOD还可避免暴露在O2-中的细菌被杀灭。早期认为羟自由基只与电离辐射有关,但实际上可能也是由呼吸生物系统产生的,这是一个发人深省的想法。由于这个想法,对SOD、过氧化氢酶和氧化物酶产生了新的认识,通过它们的联合作用,使得O2- 和H2O2 的稳态浓度微乎其微,从而抑制了Haber & Weiss反应,才使得有氧生活成为可能。
